J Pharm Pharmaceut Sci (www.cspscanada.org) 10(3): 350-357, 2007
Reversal of Multidrug Resistance by Transferrin-Conjugated Liposomes Co-encapsulating Doxorubicin and Verapamil
Jun Wu1, Yanhui Lu1, Alice Lee1,2, Xiaogang Pan1,2, Xiaojuan Yang1,2, Xiaobin Zhao1,3, Robert J. Lee1,2,3*
1 Division of Pharmaceutics, College of Pharmacy; 2 NSF Nanoscale Science and Engineering Center (NSEC)-Center for Affordable Nanoengineering of Polymeric Biomedical Devices (CANPBD); 3 NCI Comprehensive Cancer Center (CCC), The Ohio State University, USA
Received Feb 29, 2007; Revised July 10, 2007; Accepted July 20, 2007; Published July 21, 2007
Corresponding Author: Robert J. Lee, Ph.D., College of Pharmacy, The Ohio State University Columbus, Phone: (614) 292-4172, Email: lee.1339@osu.edu
ABSTRACT - PURPOSE. Liposomes co-encapsulating doxorubicin (DOX) and verapamil (VER), and conjugated to transferrin (Tf-L-DOX/VER) were synthesized and evaluated in K562 leukemia cells. The design of this formulation was aimed at selective targeting of tumor cells, reducing cardiotoxicity of DOX and VER, as well as overcoming P-glycoprotein (Pgp)-mediated multidrug resistance (MDR) phenotype. METHODS. The liposomes were prepared by polycarbonate membrane extrusion, followed by pH-gradient driven remote loading and Tf conjugation. Kinetics of in vitro release of DOX and VER from liposomes was determined by measuring changes in the concentration of encapsulated drugs. Uptake of Tf-conjugated liposomes by K562 cells was evaluated by fluorescence microscopy and by fluorometry. Cytotoxicities of various formulations of DOX were determined by a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolum bromide (MTT) assay. RESULTS. Efficiencies for liposomal loading of DOX and VER were 95% and 70%, respectively. The mean particle diameter for the liposomes was approximately 110nm. Rates of release for DOX and VER were similar in singly-loaded and co-loaded liposomes. Tf-L-DOX/VER showed efficient uptake by the TfR+ K562 cells. In DOX-resistant K562 cells (K562/DOX), Tf-L-DOX/VER showed 5.2 and 2.8 times greater cytotoxicity (IC50 = 4.18 μM) than non-targeted liposomes (L-DOX/VER) (IC50 = 21.7 μM) and Tf-targeted liposomes loaded with DOX alone (Tf-L-DOX) (IC50 = 11.5 μM), respectively. CONCLUSIONS. The combination of TfR targeting and co-encapsulation of DOX and VER was highly effective in overcoming drug resistance in K562 leukemia cells.
Introduction
Multidrug resistance (MDR) is a major obstacle to the successful application of cancer chemotherapy [1,2]. MDR is frequently associated with upregulation of the permeability-glycoprotein (Pgp), an ATP-dependent efflux pump that reduces intracellular drug concentration [3]. To overcome MDR, a variety of approaches have been developed, including the use of cytotoxic drugs in combination with a Pgp inhibitor [4-8]. Verapamil (VER), a calcium channel antagonist, has been shown to have Pgp inhibitory activity. However, the dose of VER required for Pgp blockade (2-6μM) is significantly higher than the current clinical dose for arrhythmia and may induce cardiotoxicity [3, 9, 10]. Liposomal delivery of VER may reduce its cardiotoxicity by reducing free VER concentration in the systemic circulation.
Liposomes are phospholipid bilayer vesicles. Compared to free doxorubicin (DOX), liposomal DOX exhibit prolonged systemic circulation time, reduced cardiotoxicity, increased solid tumor accumulation due to the enhanced permeability and retention (EPR) effect, and increased therapeutic efficacy in murine tumor models [11-13]. In addition, liposomal delivery has been reported to reverse the MDR phenotype [14-16]. However, other reports seem to indicate that liposomal delivery per se may have limited effectiveness in reversing MDR [17-19]. Recently, a novel liposomal formulation containing co-loaded DOX and VER was synthesized and shown to overcome MDR [20-21]. Targeted liposomal delivery has been shown previously to increase tumor cell-selective cytotoxicity and to facilitate Pgp bypass and MDR reversal [22].
Human transferrin (Tf, M.W. ~ 80kDa) is a nonheme iron-binding glycoprotein with high affinity for the Tf receptor (TfR). The TfR is frequently over-expressed on tumor and leukemia cells and undergoes internalization via TfR-mediated endocytosis [23, 24]. TfR has been studied extensively as a marker for tumor cell targeted drug delivery [25-27]. For example, Tf-conjugated liposomal DOX has been shown to selectively target tumor cells [26,28].
In this report, liposomes co-loaded with DOX and VER are further conjugated to Tf to provide a mechanism for tumor cell-selective targeting. The liposomes were evaluated for in vitro uptake and cytotoxicity in regular as well as in DOX-resistant K562 cell lines (K562/DOX).
Materials and METHODS
Materials
Egg phosphatidylcholine (EPC), cholesterol (CHOL), distearoyl phosphatidylethanolamine-N-monomethoxy-polyethylene glycol (M.W. 2,000) (mPEG-DSPE) and distearoyl............. phosphatidylethanolamine-N-maleimide polyethylene glycol 2000 (MAL-PEG-DSPE) were purchased from Avanti Polar Lipids Inc. (Alabaster, AL, USA). Human holo-transferrin (Tf), 2-iminothiolane (Traut's reagent), 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl tetrazolium bromide (MTT), DOX and VER were purchased from Sigma Chemical Co. (St. Louis, MO, USA). All chemicals used were of analytical or HPLC grade and were used without further purification.
Cell culture
K562 is a chronic myelogenous leukemia cell line with amplified TfR expression. DOX-resistant K562 was developed from regular K562 cells by culturing of these cells in the presence of increasing concentrations of DOX. DOX-resistant and regular K562 cells were kindly provided by Dr. Duxin Sun in the Division of Pharmaceutics, College of Pharmacy, The Ohio State University (Columbus, OH). DOX-resistant cells were exposed to 0.1μM DOX once per month to maintain its drug-resistant phenotype. The cells were cultured in RPMI 1640 media (Life Technologies Inc., Bethesda, MD, USA), supplemented with 50μg/mL penicillin, 50μg/mL streptomycin, and 10% fetal bovine serum (FBS) and were maintained at 37°C in a humidified atmosphere containing 5% CO2.
Liposome preparation
Liposomes were prepared by polycarbonate membrane extrusion followed by remote loading of DOX [29]. The lipid composition of liposomes was EPC/CHOL/mPEG-DSPE at a molar ratio of 55:40:5. For Tf-targeted liposomes, the composition was EPC/CHOL/mPEG-DSPE/MAL-PEG-DSPE at molar ratios of 55:40:4:1. Lipids were dissolved in CHCl3 and dried in a round-bottom flask on a rotary evaporator and then under vacuum. The resulting lipid film was hydrated with (NH4)2SO4 solution (250 mM). The lipid suspension was then extruded 5 times each through a 200-nm and a 100-nm pore size polycarbonate membrane on a nitrogen-driven LipexTM lipid extruder from Northern Lipids Inc. The liposomes were then subjected to diafiltration using a Spectrum Microkros hollow fiber cartridge to replace (NH4)2SO4 outside the liposomes with phosphate buffered saline (PBS, pH 7.4). For drug loading, DOX and VER were dissolved in a small volume of H2O and added to the liposomes. The liposomes were incubated for 30 min at 40°C and then purified by size exclusion chromatography on a Sepharose CL-4B column. Liposome size distribution was determined by dynamic light scattering on a NICOMP Submicron Particle Sizer Model 370 (NICOMP, Santa Barbara, CA, USA). To analyze DOX concentration, the liposomes were lysed with ethanol. DOX concentration in the lysate was determined by absorption at 480 nm on a Shimadzu UV-visible spectrophotometer based on the molar extinction coefficient of DOX. VER concentration was measured by HPLC using a reverse-phase AllTech RP C18 column (4.1mm×300 mm, 10µm particle size) using an isocratic elution program with a solvent system of CH3CN/20 mM phosphate buffer (pH 4.0) (50:50, V/V) on a Beckman Gold HPLC system equipped with a UV detector at 210 nm (flow rate 1mL/min).
Conjugation of Tf to liposomes
Tf-conjugated liposomes were prepared by coupling thiolated Tf to drug-loaded liposomes containing 1 mole% MAL-PEG-DSPE [30,31]. To thiolate the Tf, Traut's reagent was reacted with Tf at a molar ratio of 10:1 for 1h in HEPES-buffered saline (HBS, pH 8.0) at room temperature. The mixture was passed through a PD-10 column, eluted with HBS (pH 6.5), to remove unreacted Traut's reagent. The Tf-SH fraction was collected in the void volume and added to the liposomes (containing 1 mole% MAL-PEG-DSPE) at the ratio of 20 µg Tf conjugate per µmol total lipid and allowed to react at room temperature for 2h. Tf-conjugated liposomes were purified by size exclusion chromatography on a Sepharose CL-4B column to remove free Tf. The liposomes were collected in the void volume fractions.
In vitro release of DOX and VER from liposomes
Rates of drug release from liposomes were determined by measuring retention of the drugs. Liposomes (100 µL) were added to 900µL of PBS (pH 7.4), human plasma, or cell culture medium (RPMI-1640) containing 10% FBS and were incubated at 37°C. Free and liposome encapsulated DOX were separated by a Sepharose CL-4B column, as described above. For DOX concentration analysis, liposomes were lysed by ethanol. At various time points (0.5, 1, 2, 4, 8, 16, 24, 48, and 72 h), DOX concentration was determined based on its fluorescence by fluorometry, using excitation at 485 nm and emission at 550 nm. VER concentration was determined by HPLC, as described above. The percentages of drug released were then calculated from percentages of drug remaining in the liposome fractions.
Cellular uptake of Tf-conjugated liposomes
Uptake of Tf-conjugated liposomes by K562 cells was evaluated by fluorescence microscopy. Tf-conjugated liposomes encapsulating DOX (Tf-L-DOX) were prepared as described above. For uptake experiments, K562 cells were incubated for 2 h at 37°C with Tf-L-DOX. The same experiment was repeated using media containing 2 mg/mL Tf to determine the effect of Tf receptor blockade. At the end of incubation, the cells were washed three times with cold PBS and photographed in fluorescence and phase contrast modes on a Nikon Eclipse 800 fluorescence microscope.
Cytotoxicity analysis
Cytotoxicities of various formulations of DOX were determined by an MTT assay, as described previously with minor modifications [20]. K562 and K562/DOX cells were transferred to 96-well tissue culture plates at 5,000 cells per well 24 h prior to drug addition. The culture medium was replaced with the medium containing serial dilutions of various drug formulations, including free DOX, non-targeted liposomal DOX (L-DOX), non-targeted liposomes co-loaded with DOX and VER (L-DOX/VER), Tf-targeted liposomal DOX (Tf-L-DOX), Tf-targeted liposomes co-loaded with DOX and VER (Tf-L-DOX/VER), a mixture of free DOX and VER (DOX/VER), a mixture of Tf-L-DOX/VER and free Tf, and a mixture of Tf-L-DOX and free Tf. Following 4 h incubation at 37°C, the cells were washed twice with PBS and cultured in fresh medium for an addition 72 h. Then, 20 µL of MTT stock solution (5 mg/mL) was added to each well and the plate was incubated for 4 h at 37°C. Media were then removed and dimethyl sulfoxide (DMSO) (100 µL) was added to dissolve the blue formazan crystals converted from MTT by live cells. Cell viability was assessed by absorbance at 570nm measured on a Bio-Rad microtiter plate reader.
Analysis of intracellular DOX
The fluorescence of DOX was used to determine uptake of DOX by leukemia cells. K562/DOX cells were incubated with various formulations of DOX for 1, 4, 8, 16, 24, 48 and 72 h at 37°C. After incubation, cells were washed with cold PBS and then lysed in ethanol and 20% sodium dodecyl sulfate (SDS). DOX concentration was then determined based on its fluorescence, as described above.
Statistical analysis
Statistical significance of differences among various DOX formulations were determined using Student's t-test and significant difference was defined as p < 0.05.
Results
Characterization of Tf-conjugated liposomes
Liposomes were prepared by polycarbonate membrane extrusion and the drugs were incorporated by remote loading. DOX and VER incorporation efficiencies were found to be 96.3±4.0% and 73.7± 3.6%, respectively. The mean diameter of a typical liposome preparation was 110 nm. Tf-targeted liposomes were stable for at least 4 weeks at 4°C in PBS (pH 7.4) without any detectable leakage of DOX and VER or reduction in binding to Tf-expressing cells.
Release of DOX and VER from liposomes
As shown in Figure 1, both liposome formulations, Tf-L-DOX/VER and Tf-L-DOX, showed minimal release (25%) of DOX in PBS (pH 7.4) after 96 h incubation at 37°C (Figure 1 A&B). In cell culture medium (containing 10% FBS), the release of DOX from Tf-L-DOX/VER and Tf-L-DOX were both ~ 35% (Figure 1 A&B). In human plasma, the release of DOX from Tf-L-DOX/VER and TF-L-DOX were both ~ 45% (Figure 1 A&B). The presence of VER in Tf-L-DOX/VER had no significant effect on the rate of DOX release. Stability of VER encapsulation in liposomes was also investigated. The results showed that the rate of release of VER from Tf-L-DOX/VER was similar to that of DOX in various media at 37°C (Figure 1C).
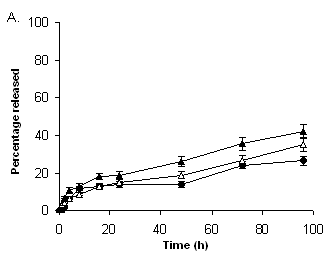
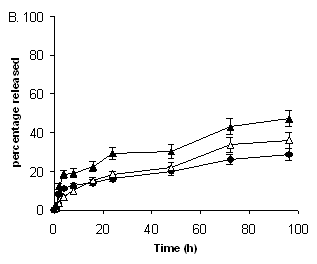
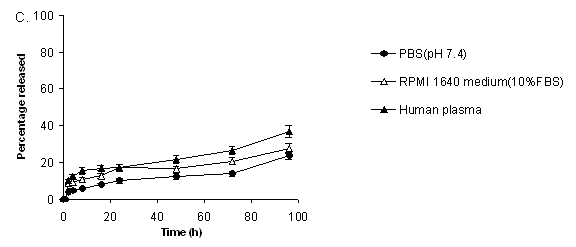
Figure 1. Kinetics of of DOX and VER release from Tf-L-DOX/VER or Tf-L-DOX in various media. (A) DOX release from Tf-L-DOX/VER; (B) DOX release from Tf-L-DOX; (C) VER release from Tf-L-DOX/VER. Error bar = 1 standard deviation (n=3).
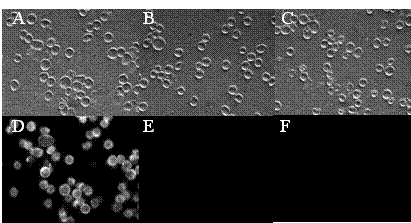
Figure 2. Uptake of Tf-L-DOX and L-DOX by K562 cells. K562 cells were treated with Tf-L-DOX, L-DOX, or Tf-L-DOX plus 2 mg/mL free Tf, and photographed in both fluorescence (upper panels) and phase-contrast (lower panels) modes on a Nikon Eclipse 800 fluorescence microscope. All the samples were incubated for 4 h at 37°C. (A) and (D): cells treated with Tf-L-DOX; (B) and (E): cells treated with L-DOX; (C) and (F): cells treated with Tf-L-DOX plus 2 mg/mL free Tf.
Cellular uptake and cytotoxicity of Tf-L-DOX/VER
Uptake of TfR-targeted liposomal DOX and VER by K562 cells was analyzed by fluorescence microscopy, as shown in Figure 2. TfR-targeted liposomes showed much greater cellular uptake and cellular internalization than non-targeted control liposomes and the uptake was blocked by the addition of 2 mg/mL free Tf. These results indicated that these liposomes have efficient interactions with the TfR on K562 cells.
Cytotoxicity of various DOX formulations was determined in K562 and K562/DOX cells using an MTT assay. IC50 values of the various formulations in K562/DOX cells, indicated the order of Tf-L-DOX/VER~DOX/VER < Tf-L-DOX < Tf-L-DOX/VER + free Tf < L-DOX/VER < Tf-L-DOX + free Tf ~ L-DOX (Table 1). The results showed that Tf-L-DOX/VER had approximately 5 times lower IC50 value than that of non-targeted L-DOX/VER. The difference of cytotoxicity between targeted and non-targeted liposomes was diminished in the presence of 2 mg/mL free Tf, suggesting TfR dependence of the cytotoxicity exhibited by the Tf-L-DOX/VER. In contrast, liposomes encapsulating DOX alone showed only minimal cytotoxicity, indicating stable sequestration of DOX by liposomal encapsulation. Time-dependent uptake of DOX in various formulations was determined by cell-associated DOX fluorescence. As shown in Figure 3, efficiencies of DOX uptake follow the order of L-DOX<DOX<L-DOX/VER<Tf-L-DOX<Tf-L-DOX/VER<DOX/VER.
Discussion
In this study, a novel Tf-conjugated liposomal formulation containing co-loaded DOX and VER was synthesized and evaluated in regular and drug-resistant K562/DOX cells. The results indicated that TfR targeting, combined with co-loading of DOX and VER, produced a liposomal formulation that is highly specific for TfR expressing cells and capable of overcoming drug resistance in K562/DOX cells.
Co-delivery of therapeutic agents in liposomes has been proposed as a mechanism to improve therapeutic synergism [30, 31]. Since both DOX and VER are weakly basic drugs, it is possible to load both agents into liposomes by remote loading via a transmembrane pH gradient. Liposomes co-loaded with DOX and VER have recently been synthesized and shown to overcome drug resistance very effectively [20]. Targeted liposomal delivery has also been shown to facilitate bypassing of Pgp-mediated multidrug resistance in cancer.
Table 1. Cytotoxicity of various DOX formulations on K562 and K562/DOX cells
|
IC50(μM) |
Reversal efficiency a) |
Resistance factor b) |
|
|
K562 |
K562/DOX |
|
|
Tf-L-DOX/VER |
0.26±0.04 |
4.18±0.37 |
5.61 |
52 |
Tf-L-DOX |
0.21±0.03 |
11.4±1.7 |
2.04 |
145 |
Tf-L-DOX/VER + free Tf |
0.96±0.22 |
19.6±2.2 |
1.19 |
249 |
Tf-L-DOX + free Tf |
1.29±0.23 |
>150 |
<0.16 |
>1898 |
L-DOX |
1.55±0.26 |
>150 |
<0.16 |
>1898 |
L-DOX/VER |
0.89±0.15 |
21.7±3.0 |
0.93 |
274 |
DOX/VER |
0.11±0.02 |
3.92±0.50 |
5.98 |
49 |
DOX |
0.079±0.016 |
23.4±3.4 |
1 |
296 |
a) Reversal efficiencies were calculated from the ratios between the mean IC50 values of free DOX and that of various DOX formulations in K562/DOX cells. b) Resistance factors were calculated from the ratios between IC50 values of various DOX formulations in K562/DOX cells and that of free DOX in K562 sensitive cells. Data are shown as mean± standard deviation (n=3).
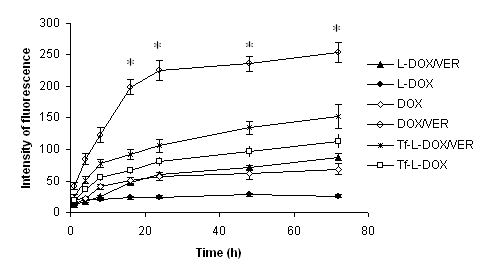
Figure 3. Accumulation of various DOX formulations in K562/DOX cells. Cells were incubated in RPMI 1640 medium containing various DOX formulations. Cell-associated DOX was then determined based on DOX fluorescence (excitation at 485nm, emission at 550nm) by quantitative fluorometry. Error bar equals 1 standard deviation (n=3). * indicates significant difference versus Tf-L-DOX/VER or p<0.05.
Our approach combines liposomal co-delivery of DOX and VER and TfR-targeting, thus is likely to have the combined advantages of tumor cell selectivity and greater reversal efficiency.
It is important to note that liposomal DOX and free DOX have very difference kinetic properties, both in terms of in vitro cellular uptake, which is diffusion driven, and in vivo plasma residence time, which is determined by the mechanism of clearance. L-DOX, depending on the liposome composition, has a circulation half-life of hours to days, compared to a half-life of minutes for free DOX. Because of this difference, direct comparison of IC50 values for L-DOX and free DOX using equal exposure time in a cytotoxicity assay is not possible, since the comparison would have no relevance to in vivo therapeutic efficacy. Comparison of efficacy of liposomal drug and free drug can only be carried out using in vivo models. However, it is possible to carry out in vitro comparisons among the various liposomal formulations that exhibit similar in vitro (diffusion) and in vivo (clearance) kinetic properties. Therefore, remaining discussions of data will mostly focus on comparisons among liposomal formulations.
In fact, our in vitro cytotoxicity studies demonstrated that TfR targeting played a pivotal role in reversing MDR. Tf-L-DOX/VER and Tf-L-DOX exhibited higher reversal efficiencies than L-DOX/VER and L-DOX, respectively. The IC50 value of L-DOX was much higher than DOX. This is because DOX is sequestered inside the liposomes. However, L-DOX has actually shown greater antitumor activity than free DOX in preclinical studies since its clearance is mediated by the reticuloendothelial system (RES), has a much longer systemic circulation half-life, and exhibits pharmacokinetics that is fundamentally different from that of free DOX. The use of equal exposure time for L-DOX and free DOX in in vitro cytotoxicity studies creates an artificial condition strongly favoring free DOX.
Our data suggest cytotoxicity depends on both cellular uptake and the mechanism of uptake. For L-DOX and L-DOX/VER, the low cytotoxicity on K562/DOX cells could be attributed to low cellular uptake. Meanwhile, Tf-L-DOX/VER was almost as cytotoxic as DOX/VER in K562/DOX cells. However, overall DOX cellular uptake of Tf-L-DOX/VER was actually lower than that of DOX/VER within 72 h. This suggested that the mechanism of cellular entry (receptor-mediated endocytosis for Tf-liposomes versus passive diffusion from free drug) is an important determinant for cytotoxicity.
conclusions
TfR-targeted liposomal delivery of co-encapsulated DOX and VER was shown to be an effective strategy for selective targeting and reversing of drug resistance in K562 cells. Further in vivo studies are warranted to investigate the efficacy and toxicity of this novel formulation.
Acknowledgements
We thank Dr. Duxin Sun for providing DOX-resistant and sensitive K562 cells. This work was supported in part by NSF grant EEC-0425626
References
| [1] | Klohs W.D., Steinkampf R.W., The effect of lysosomotropic agents and secretory inhibitors on anthracycline retention and activity in multiple drug-resistant cells. Mol Pharmacol, 34: 180-185, 1988. |
| [2] | Sehested M., Skovsgaard T., Roed H., The carboxylic ionophore monensin inhibits active drug efflux and modulates in vitro resistance in daunorubicin resistant Ehrlich ascites tumor cells. Biochem Pharmacol, 37: 3305-3310, 1988. |
| [3] | Krishna R., Mayer L.D., Multidrug resistance (MDR) in cancer Mechanisms, reversal using modulators of MDR and the role of MDR modulators in influencing the pharmacokinetics of anticancer drugs, Eur J Pharm Sci, 11: 265-283, 2000. |
| [4] | Vezmar M., Georges E., Reversal of MDR-mediated doxorubicin resistance with quinolone-based drugs. Biochem Pharmacol, 59: 1245-1252, 2000. |
| [5] | Belpomme D., Gauthier S., Pujade-Lauraine E., Verapamil increases the survival of patients with anthracycline-resistant metastatic breast carcinoma. Ann Oncol, 11: 1417-1476, 2000. |
| [6] | Aszalos A., Ladanyi A., Bocsi J., Szende B., Induction of apoptosis in MDR1 expressing cells by daunorubicin with combinations of suboptimal concentrations of P-glycoprotein modulators. Cancer Lett, 167: 157-162, 2001. |
| [7] | Merlin J., Bour-Dill C., Marchal S., Ramacci C., Poullain M., Giroux B., Modulation of daunorubicin cellular resistance by combination of P-glycoprotein blockers acting on drug efflux and intracellular drug sequestration in Golgi vesicles. Cytometry, 41: 62-72, 2000. |
| [8] | Volm M., Multidrug resistance and its reversal. Anticancer Res, 18: 2905-2918, 1998. |
| [9] | Mross K, Hamm K., Hossfeld D.K., Effects of verapamil on the pharmacokinetics and metabolism of epirubicin. Cancer Chemother Pharmacol, 31: 369-375, 1993. |
| [10] | Pennock G.D., Dalton W.S., Roeske W.R., Appleton C.P., K. Mosley, P. Plezia, T.P. Miller, S.E. Salmon, Systemic toxic effects associated with high-dose verapamil infusion and chemotherapy administration. J Natl Cancer Inst, 83: 105-110, 1991. |
| [11] | Harrington K.J., Mohammadtaghi S., Uster P.S, Glass D., Peters A.M., Vile R.G., Stewart J.S., Effective targeting of solid tumors in patients with locally advanced cancers by radiolabeled pegylated liposomes. Clin Cancer Res, 7: 243-254, 2001. |
| [12] | Allen T.M., Long-circulation (sterically stabilized) liposomes for targeted drug delivery. Trends Pharmacol Sci, 15: 215-220, 1994. |
| [13] | Gabizon A.A., Selective tumor localization and improved therapeutic index of anthracyclines encapsulated in long-circulating liposomes. Cancer Res, 52: 891-896, 1992. |
| [14] | Mayer L.D., Shabbits J.A., The role for liposomal drug delivery in molecular and pharmacological strategies to overcome multidrug resistance. Cancer Metastasis Rev, 20: 87-93, 2001. |
| [15] | Thierry A.R., Vige D., Coughlin S.S., Belli J.A., Dritschilo A., Rahman A., Modulation of doxorubicin resistance in multidrug-resistant cells by liposomes. FASEB J, 7: 572-579, 1993. |
| [16] | Warren L., Jardillier J.C., Malarska A., Akeli M.G., Increased accumulation of drugs in multidrug-resistant cells induced by liposomes. Cancer Res, 52: 3241-3245, 1992. |
| [17] | Hu Y.P., Henry-Toulme N., Robert J., Failure of liposomal encapsulation of doxorubicin to circumvent multidrug resistance in an in vitro model of rat glioblastoma cells. Eur J Cancer, 31A: 389-394, 1995. |
| [18] | Krishna R., St Louis M., Mayer L.D., Increased intracellular drug accumulation and complete chemosensitization achieved in multidrug-resistant solid tumors by co-administering valspodar (PSC 833) with sterically stabilized liposomal doxorubicin. Int J Cancer, 85: 131-141, 2000. |
| [19] | Rivera E., Valero V., Esteva F.J., Syrewicz L., Cristofanilli M., Rahman Z., Booser D.J., Hortobagyi G.N., Lack of activity of stealth liposomal doxorubicin in the treatment of patients with anthracycline-resistant breast cancer. Cancer Chemother Pharmacol, 49: 299-302, 2002. |
| [20] | Wang J.C., Goh B., Lu W.L., Zhang Q., Chang A., Liu X.Y., Tan T. MC., Lee H.S., In vitro cytotoxicity of stealth liposomes co-encapsulating doxorubicin and verapamil on doxorubicin-resistant tumor cells. Biol Pharm Bull,28: 822-828, 2005. |
| [21] | Wong H.L., Bendayan R., Rauth A.M., Wu X.Y., Development of solid lipid nanoparticles containing ionically complexed chemotherapeutic drugs and chemosensitizers. J Pharm Sci, 93: 1993-2008, 2004. |
| [22] | Huwyler J., Cerletti A., Fricker G., Eberle A.N., Drewe J., By-passing of P-glycoprotein using immunoliposomes. J Drug Target, 10: 73-79, 2002. |
| [23] | Huebers H.A., Finch C.A., The physiology of transferrin and transferrin receptors. Physiol Rev, 67: 520-582, 1987. |
| [24] | Singh M., Transferrin as a targeting ligand for liposomes and anticancer drugs, Curr Pharm Des, 5: 443-451, 1999. |
| [25] | Derycke A.S., Kamuhabwa A., Gijsens A., Roskams T., De Vos D., Kasran A., J. Huwyler, L. Missiaen. P.A. de Witte, Transferrin-conjugated liposome targeting of photosensitizer A1PcS4 to rat bladder carcinoma cells. J Natl Cancer Inst, 96: 1620-1630, 2004. |
| [26] | Qian Z.M., Li H., Sun H., Ho K., Targeted drug delivery via the transferrin receptor-mediated endocytosis pathway. Pharmacol Rev, 54: 561-587, 2002. |
| [27] | Linuma H., Maruyama K., Okinaga K., Sasaki K., Sekine T., Ishida O., Ogiwara N., Johkura K., Yonemura Y., Intracellular targeting therapy of cisplatin-encapsulated transferrin-polyethylene glycol liposome on peritoneal dissemination of gastric cancer. Int J Cancer, 99: 130-137, 2002. |
| [28] | Evavarone D.A., Yu X., Bellamkonda R.V., Targeted drug delivery to C6 glioma by transferrin-coupled liposomes. J Biomed Mater Res, 51: 10-14, 2000. |
| [29] | Haran G., Cohen R., Bar L.K., Barenholz Y., Transmembrane ammonium sulfate gradients in liposomes produce efficient and stable entrapment of amphipathic weak bases. Biochim Biophys Acta, 1151: 201-215, 1993. |
| [30] | Provoda C.J.,. Stier E.M., Lee K.D., Tumor cell killing enabled by listeriolysin O-liposome-mediated delivery of the protein toxin gelonin. J Biol Chem, 278: 35102-35108, 2003. |
| [31] | Abraham S.A., McKenzie C., Masin D., Ng R., Harasym T.O., Mayer L.D., and Bally M.B., In vitro and in vivo characterization of doxorubicin and vincristine coencapsulated within liposomes through use of transition metal ion complexation and pH gradient loading, Clin Cancer Res, 10, 728-738, 2004. |
Published by the Canadian Society for Pharmaceutical Sciences.
Copyright © 1998 by the Canadian Society for Pharmaceutical Sciences.